Urheberrechte
Dieser Artikel wurde der "Wikipedia" entnommen und steht unter dieser Nutzungslizenz.
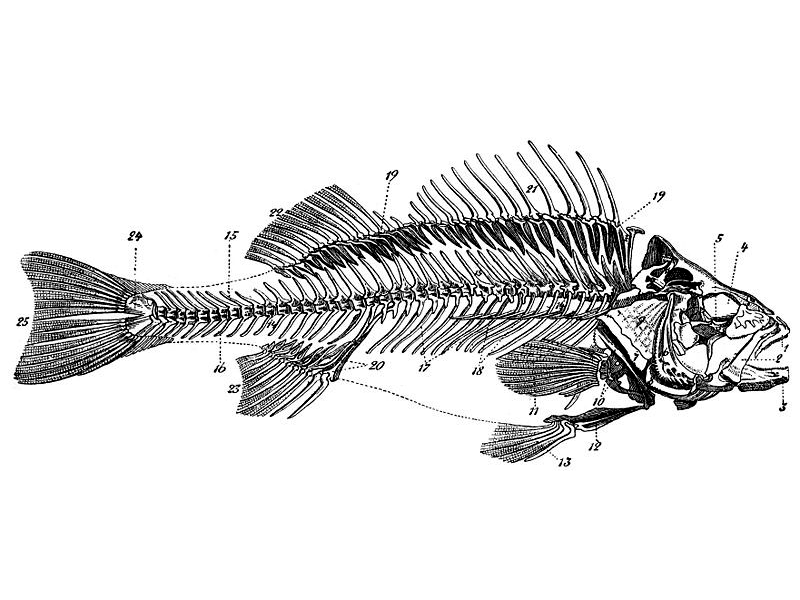
Das Skelett besteht aus Knochen, bei primitiven (aber auch abgeleiteteren) Arten teilweise noch aus Knorpel.
Am Schädel befindet sich das Kiemenskelett aus sieben Kiemenbögen, von denen der vordere zum Unterkiefer umgebildet ist.
In der Rumpfregion tragen die Wirbel Rippen, über die ganze Länge Dornfortsätze auf der Rückenseite, im Schwanzbereich auch auf der Bauchseite.
In den Muskelscheiden von Knochenfischen befinden sich häufig Gräten aus verknöchertem Bindegewebe.
Zur Steuerung und Fortbewegung dienen die Flossen, die bis auf die Fettflosse (bei Lachsartigen, Salmlerartigen, Welsartigen) mit knöchernen Flossenstrahlen (Radien, aus Schuppenreihen entstanden) verstärkt sind. Brust- und Bauchflossen sind paarig und gelenken am Schultergürtel beziehungsweise am Becken. Die unpaarigen Rücken-, After- und Schwanzflosse(n) sind über Flossenträger (Radialia) mit der Wirbelsäule verbunden.
Die erwähnte Fettflosse hat kein Skelett, kann aber hornartig verstärkt sein.
Die Muskulatur der Knochenfische wird in Rumpf- und Flossenmuskulatur unterteilt. Die Rumpfmuskulatur besteht
aus Einzelsegmenten, deren Anzahl meist der der Wirbel entspricht.
Der Mondfisch hat nur 17 Wirbel, der Schnepfenaal (Nemichthys scolopaceus) über 600. Am Schädel setzt naturgemäß eine große Zahl von Muskeln für Nahrungserwerb und Atmung an. Sie sind segmental in Myomere gegliedert.
Die Scheidewände zwischen den Myomeren heißen Myosepten. Die Myomere verlaufen W-förmig wenn man sie von der Seite betrachtet. Bei manchen Arten (vor allem Pflanzenfresser) entwickelt siche ein muskelstarker Kaumagen.
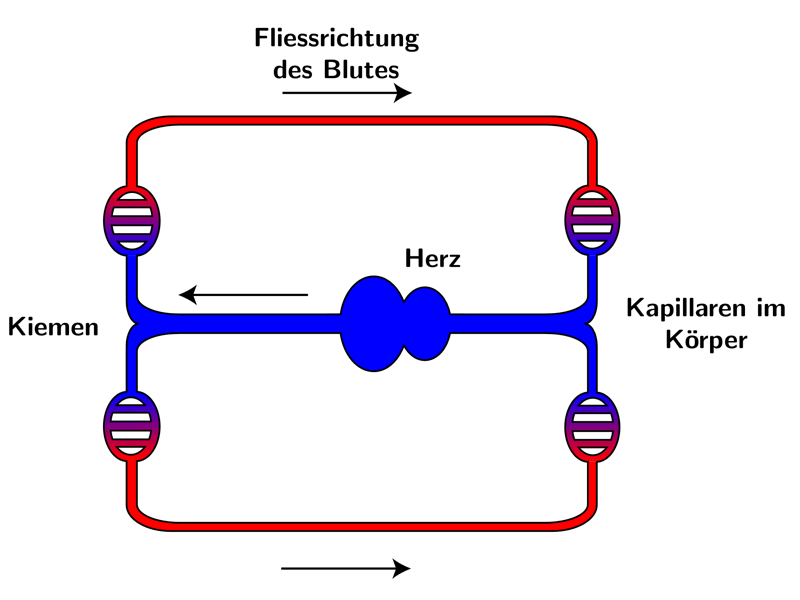
Knochenfische haben einen geschlossenen Blutkreislauf mit einem nahe an den Kiemen gelegenen, einfachen Herzen, das aus einem Vorhof und einer Herzkammer besteht.
Es pumpt venöses Blut direkt in die Kiemen, die aus zahlreichen, mit Kapillaren durchzogenen Blättchen, feinen Hautausstülpungen, bestehen.
Das nebenstehende Bild zeigt eine schematische Darstellung des Blutkreislaufs:
Rot = Sauerstoffreiches Blut
Blau = Sauerstoffarmes Blut
Die Kiemen nehmen über ihre große Oberfläche gelösten Sauerstoff aus dem Wasser auf und geben ihn an das Blut weiter. Die sehr hohe Effektivität der Sauerstoffaufnahme über die Kiemen (bis zu 70 % des verfügbaren Sauerstoffs) kann auf das Prinzip des Gegenstrom-Austausches in den Kiemen und die sehr hohe Affinität von einigen Knochenfisch hämoglobinen zu Sauerstoff zurückgeführt werden.
Die Kiemen sind in der Kiemenhöhle durch den knöchernen Kiemendeckel (Operculum) geschützt. Durch die Absenkung des Mundbodens und das gleichzeitige Abheben des Kiemendeckels strömt Atemwasser durch das Maul ein, während dies am Kiemendeckel durch eine weiche Membran verhindert wird. Beim Auspressen schließen sich im vorderen Maulbereich zwei Klappen (Valven) und das Wasser wird durch die Kiemendeckelspalte nach hinten ausgepresst; zu beachten ist, dass diese beiden Vorgänge fast synchron ablaufen, so dass ständig Wasser durch die Kiemenspalten fließen kann.
Das Blut dient auch dem Transport der in den inkretorischen Drüsen gebildeten Hormone – welche (beide!) schon prinzipiell denen des Menschen gleichen. Bei einigen Knochenfischen (beispielsweise Aalen) ist die Kiemenhöhle nur durch einen kleinen Spalt nach außen geöffnet, wodurch die Kiemen auch auf dem Trockenen eine gewisse Zeit feucht und damit funktionsfähig bleiben.
Einige mehr oder weniger amphibisch lebende Fischarten verfügen über weitere Atmungsorgane: Schlammspringer können Luftsauerstoff durch Hautatmung aufnehmen. Verbreiteter ist die Darmatmung, beispielsweise beim Schlammpeitzger, dabei wird Luft geschluckt, der Gasaustausch findet an den Wänden des stark durchbluteten Vorderdarms statt.
Einige Fische (Labyrinthfische) atmen auch Luft, indem sie Ausstülpungen im Kiemenbereich zur Aufnahme von Sauerstoff aus der Luft nutzen. Fischen, deren Schwimmblase noch mit dem Darm verbunden ist, wie den Knochenhechten, dient auch die Schwimmblase zur Luftatmung. Bei Flösselhechten und Lungenfischen haben sich diese Darmausstülpungen bereits zu Lungen entwickelt

")
Schwimmblase einer Rotfeder
Bei den meisten Knochenfischarten hat die Schwimmblase die Funktion, das spezifische Gewicht (die Dichte) zu regulieren, so dass sie ohne Kraftanstrengung im Wasser schweben können oder nur sehr langsam zu Boden sinken.
Hervorgegangen ist sie aus einer Ausstülpung an der Oberseite des Vorderdarms; sie kann, wie bei den Karpfen, noch mit ihm verbunden sein (physostom) oder, wie bei den Barschen, vom Darm getrennte Kammern bilden (physoklist). Zur Regulierung des spezifischen Gewichts werden Gase (Sauerstoff, Kohlendioxid, Stickstoff) aus dem Blut über die so genannte Gasdrüse oder den Verbindungsgang zum Darm, den Ductus pneumaticus, in die Schwimmblase abgegeben.
Zur Volumenverminderung der Schwimmblase gelangt das Gas entweder über den ductus pneumaticus in den Vorderdarm (und wird ausgespien) oder über einen stark durchbluteten Bereich in der Schwimmblasenwand, das Oval, wieder zurück ins Blut. Knorpelfischen, aber auch bodenlebenden oder besonders gut schwimmenden Knochenfischen fehlt die Schwimmblase – sie sinken zu Boden, wenn sie sich nicht fortbewegen.
Durch Knochenreduktion, Fetteinlagerung u. ä. kann aber z. B. der Mondfisch (Mola) auch ohne Schwimmblase an der Meeresoberfläche treiben.
Das Nervensystem ist einfach, das Gehirn klein, eine Großhirnrinde fehlt. Der Geruchssinn der Fische ist meist sehr ausgeprägt, Nasen- und Mundhöhle sind voneinander getrennt.
Die Gleichgewichts- und Gehörorgane bestehen aus geschlossenen, flüssigkeitsgefüllten Blasen. Bei einigen Arten (Karpfen, Salmler, Welse) sind sie durch bewegliche Knochen mit der Schwimmblase verbunden und werden als Weberscher Apparat bezeichnet. Im Aufbau ähneln sie dem Innenohr der Säugetiere, eine Hörschnecke ist jedoch nicht ausgebildet.
Die meisten Fischarten können Signale unterhalb 1 kHz wahrnehmen, einige haben ihr Hörvermögen durch spezielle Strukturen verbessert und den Hörbereich bis 5 oder mehr kHz erweitert.
Die Augen haben eine starre Linse, die auf Nahsicht fokussiert ist, aber durch einen Muskel auf Fernsicht zurückgezogen werden kann. Viele Fische sind farbtüchtig und nehmen auch ultraviolettes Licht wahr.
Besonders bei bodenlebenden Fischen ist der Tastsinn gut entwickelt. Vor allem an Lippen und Barteln befinden sich in der Oberhaut Sinneszellen, die Berührungs-, aber natürlich auch Geschmacksreize übertragen.
Als „sechsten Sinn“ verfügen Fische über Organe zur Wahrnehmung von Wasserströmungen, die Seitenlinienorgane, die sich seitlich etwa in der Mitte über einen Großteil der Körperlänge erstrecken. Sie bestehen aus einer Reihe von Hauteinstülpungen, in denen sich Sinneszellen mit Sinneshaaren befinden, die durch Strömungsveränderungen erregt werden.
Wenn diese „Seitenlinie“ fehlt (z. B. beim Hering), gibt es doch auch am Kopf solche Organe bei (fast) allen Knochenfischen.
")
Laich des Moderlieschens
Die Nieren sind ein paariges (oder unpaarig verschmolzenes) langgestrecktes Organ unterhalb der Wirbelsäule.
Bei allen anderen "Fischen" dienen die Harnleiter (oder davon abgegliederte Röhren) auch der Ausleitung der Keimzellen - aber die Echten Knochenfische haben dazu eigene Ei- und Samenleiter entwickelt.
Ihre Fortpflanzungsorgane befinden sich seitlich und oberhalb des Darms.
Die Befruchtung findet bei den meisten Arten ohne Kopulation statt.
Stattdessen wird der weibliche Rogen (die Eier) und die männliche Milch (der Samen) etwa gleichzeitig ins Wasser abgesetzt.
Der zur Fortpflanzung fähige weibliche Fisch heißt "Rogner", der fortpflanzungsfähige männliche Fisch wird "Milchner" genannt.
Dabei schwankt die Anzahl der Eier extrem: Störe legen mehrere Millionen, die brutpflegenden Stichlinge nicht mehr als hundert. Bei brutpflegenden Arten sind es häufig die Männchen, die die Pflege übernehmen. Es gibt bei den brutpflegenden Arten verschiedene Formen: Offen-, Höhlen- und Maulbrüter. Bei Offenbrütern werden die Eier vom Weibchen auf Pflanzen oder anderen festen Materialien ablegt und anschließend vom Männchen besamt.
Bei Höhlenbrütern werden die Eier an der Decke von Höhlen abgelegt. Nach dem Schlüpfen werden die Jungen noch eine Zeit lang auf Blättern oder in Gruben abgelegt (bis der Dottersack verbraucht ist) und bewacht. Danach werden die Larven von den Eltern geführt, wobei sich die Eltern entweder abwechseln oder ein Elternteil, meistens das Männchen, die Reviergrenzen bewacht. Geht ein Großteil der Brut verloren, so kann es vorkommen, dass die Eltern die wenigen überlebenden Larven fressen und schnell wieder mit einer neuen Brut beginnen.
Bei Maulbrütern nehmen die Weibchen die Eier nach der Ablage ins Maul, das Männchen schwimmt über die Eier und gibt dabei seinen Samen ab. Die Jungen der Maulbrüter sind, wenn sie das Maul verlassen, schon weit entwickelt, sie kehren nach dem Schlüpfen aber noch eine Zeit lang bei Gefahr ins Maul zurück.
Kuckucks-Fiederbartwelse haben sich darauf spezialisiert, ihre Eier zu den Eiern des Wirtsfisches abzulegen, die dann vom Weibchen mit ins Maul aufgenommen und dort "ausgebrütet" werden. Die sich schneller entwickelnden Welslarven fressen dann die Eier oder Larven des Wirtsfisches. Diskusfische sondern nach dem Schlüpfen ein Hautsekret ab, das von den Larven abgeweidet wird und als Erstnahrung dient.
Bei Labyrinthfischen bilden die Männchen häufig an der Wasseroberfläche zwischen Schwimmpflanzen ein Schaumnest und treiben dann das Weibchen zur Eiablage unter das Schaumnest. Dabei umschlingt das Männchen mit seinem Körper das Weibchen und dreht es auf den Rücken, so dass die Eier nach oben schwimmen. Nach dem Besamen wird das Weibchen vertrieben und die Brut vom Männchen allein behütet bis die Larven schlüpfen, danach kümmert sich auch das Männchen nicht weiter um die Brut.
Einige Knochenfische, besonders marine Barschartige, haben kein genetisch bestimmtes Geschlecht, es prägt sich erst durch den Kontakt mit Partnern oder durch Umweltbedingungen aus und kann mehrmals gewechselt werden.
Es gibt auch (selten) Knochenfische, die über Kopulationsorgane verfügen, wie beispielsweise die Lebendgebärenden Zahnkarpfen (Poeciliidae). Die Larven der Knochenfische entwickeln sich durch Metamorphose zu Adulttieren.
Bei der Metamorphose werden die larvalen Organe resorbiert oder abgestoßen und die vorhandenen Anlagen der Adultorgane zur Funktionsfähigkeit entwickelt.
Beide Klassen der Knochenfische, Muskelflosser und Strahlenflosser, sind bei ihrem ersten Erscheinen im Unteren Devon schon deutlich voneinander getrennt.
Der erste Quastenflosser war Eoactinistia foreyi. Die ersten Formen lebten als kleine, räuberische spindelförmige Fische im Meer. Beide Gruppen fanden aber sehr schnell neue Lebensräume im Süßwasser.
Die Muskelflosser waren während des Mittleren Devon die dominierenden Fische der Süßgewässer. Aus ihnen gingen während des Oberen Devon über Übergangsformen, wie Eusthenopteron, Panderichthys und Tiktaalik die Landwirbeltiere (Tetrapoda) hervor. Am Ende des Perm wurde die Artenzahl der Muskelflosser stark reduziert.
Fast alle Formen verschwanden mit dem Ende der Kreide. Erst im 19. Jahrhundert wurden die 6 Arten der Lungenfische und 1938 der Komoren-Quastenflosser (Latimeria chalumnae) a ls "Lebende Fossilien" entdeckt.
Ursprüngliche Formen der Strahlenflosser, die Knorpelganoiden (Chondrostei), erlebten eine rasche Radiation im Karbon, um am Ende der Trias wieder weitgehend zu verschwinden. Die Flössler (Polypteriformes) und die Störartigen (Acipenseriformes) sind die noch heute lebenden nächsten Verwandten dieser urtümlichen Knochenfische.
Eine weitere Gruppe, die basalen Taxa der Neuflosser (Neopterygii) evolvierte rasch während der Trias und verschwand zum großen Teil am Ende der Kreide. Zu diesen Fischen gehörte Leedsichthys problematicus, der größte Knochenfisch der Erdgeschichte, der möglicher weise eine Länge von bis zu 25 Metern erreicht hat.
Die nord- und mittelamerikanischen Knochenhechte (Lepisosteidae) und der nordamerikanische Kahlhecht oder Schlammfisch (Amia calva) sind Überlebende dieser zweiten Blüte der Knochenfische. Im Trias vor etwa 220 Mio. Jahren traten erstmals die Echten Knochenfische (Teleostei) auf.
Die folgend Liste zeigt die Verwandtschaft der basalen Taxa der Knochenfische bis zu den Teleostei. In Klammern
ist die Anzahl der heute lebenden Arten angegeben.
Während der mittleren Kreide gab es einen explosionsartigen Evolutionsschub, bei dem unter anderen die Protacanthopterygii entstanden.
Schon vor dem Ende der Kreide hatten die Echten Knochenfische ihre Konkurrenten weitgehend verdrängt. Eine zweite Phase rascher evolutionärer Aufspaltung in der oberen Kreide und im unteren Tertiär führte zur Entstehung der Barschartigen (Perciformes).
Nur 57 % aller heute lebenden Familien der Echten Knochenfische sind fossil überliefert. Fast 70 Familien sind ausgestorben.
Der Erfolg der Echten Knochenfische beruht zu einem großen Teil auf der Anpassung an das Leben im freien Wasser. Es gibt im Laufe der Knochenfischevolution eine Tendenz zur Verminderung der Anzahl der Wirbel, wodurch der Fischkörper steifer und strömungsgünstiger wird.
Außerdem erschlossen sich die Echten Knochenfische durch Veränderungen im Schädel (Kieferapparat!) die Aufwuchsnahrung auf Felsen und in Korallenriffen. Sie stellen heute mit über 40 Ordnungen und über 400 Familien den größten Teil der Fischarten.
Einige Taxa der Echten Knochenfische, wie die Maulstachler (Stomiiformes), die Eidechsenfischverwandten (Aulopiformes) und die Laternenfischverwandten (Myctophiformes) passten sich einem Leben in der Tiefsee an.
Verwandtschaft der Echten Knochenfische bis zu den Stachelflossern (Acanthopterygii):
Die Evolution und die phylogenetische Verwandtschaft unter den Arten, Gattungen und Familien der Stachelflosser (Acanthopterygii) ist noch weitgehend ungeklärt. Die Fülle von fast 15000 Arten erschwert die genaue Erforschung.
Dieser Artikel wurde der "Wikipedia" entnommen und steht unter dieser Nutzungslizenz.
Viele Bilder unseres Fischlexikons sind durch Creative Commons (abgekürzt CC) lizenzrechtlich geschützt. Creative Commons ist nicht der Name einer einzigen Lizenz. Die verschiedenen Lizenzen von Creative Commons weisen vielmehr große Unterschiede auf. Weitere Informationen zu diesen Lizenzen findet Ihr [hier].
Alle Artikel unseres Fischlexikons dienen ausschließlich der allgemeinen Information und erheben keinen Anspruch auf Richtigkeit und Vollständigkeit. Für die Vollständigkeit und Richtigkeit wird keine Haftung übernommen.
Alle Bilder wurden von uns digital bearbeitet und beschnitten. Weitere Infos unter "Bildrechte".